מבחן לדוגמה:
הקדמה:
ביוכימיה עוסקת בהכרת הכימיה האורגנית (החומרים אשר משמשים ליצירת והפעלת הגוף החי), ולהכרת המסלולים המטבוליים בגוף הבריא והחולה. תחילה מוצגים החומרים: פחמימות, ליפידים, חלבונים. לאחר מכן יוצגו מסלולים מטבוליים אשר משמשים ליצירת חומרים מורכבים יותר/ יציבים יותר או פירוקם לחומרים זמינים/ פעילים.
פחמימות
פחמימות מכילות קשר כפול עם חמצן בקצה או לאורך השרשרת, אם הקשר בקצה המולקולה מכונה אלזוד ואם לאורך השרשרת ולא בקצה קלטוז, בכל יתר המקומות מכילה מימן והידרוקסיל (OH).
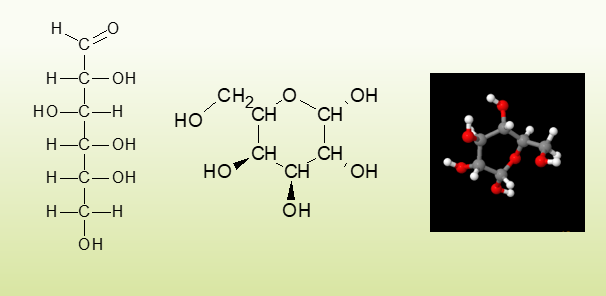
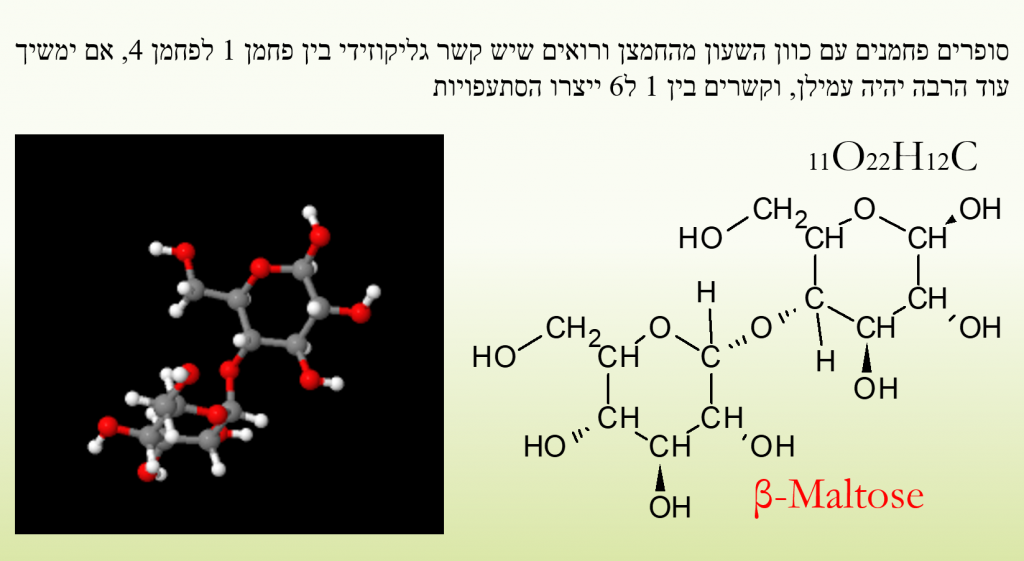
להלן איור של גלוקוז, פחמן 1 מתחבר לפחמן 5 על ידי ניתוק קשר אחד מהחמצן שבפחמן 1, ספיחת מימן, בפחמן 5 קבוצת הידרוקסיל (OH) מתפרקת משתחרר מימן והחמצן שנותר משמש לקשר גליקוזידי.
עוד תכונה של פחמימות שיש להן פחמנים קיריליים (מלשון כף יד) המשמעות היא יש למולקולה עם פחמן קירילי מולקולה המהווה תמונת ראי כמו שיש יד ימנית ויד שמאלית והן לעולם לא יהיו אותו הדבר, גוף האדם מסוגל לפרק מולקולות מסוג D, שההידרוקסיל בהן ממוקם מימין כשהחמצן למעלה(לעניות דעתי, ייתכן שבעתיד אכתוב הגדרה יותר מדעית), ולא יוכל לפרק מולקולות מסוג L, הדרך היחידה להבדיל היא ההבדל שלהם בשבירת קרני אור, חומרים כמו סוכרזית מהווים תמונת ראי של סוכר אותו אנו מסוגלים לעכל ואינם נספגים.
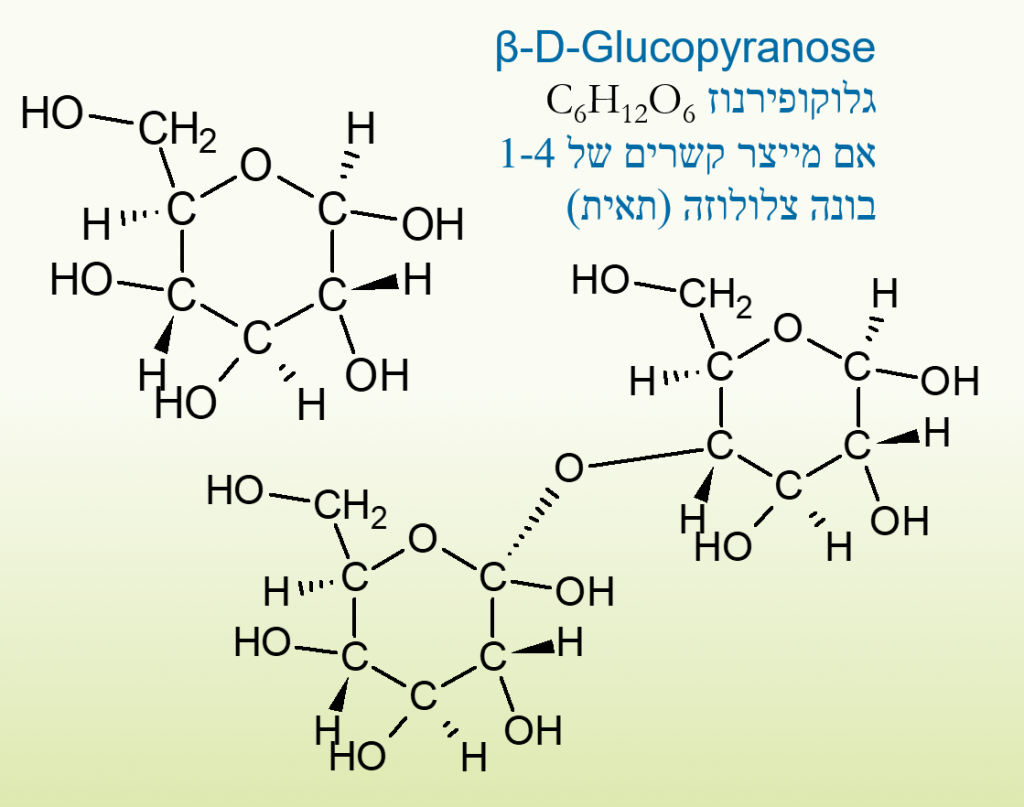
סוכר יכול ליצור דו סוכרים, כאשר נוצר קשר גליקוזידי כמו שנוצר בגלוקוז בין פחמנים 1, ו4 בדרך כלל, הקשר יכול להיקרא קשר אלפא וגם קשר בטא, בקשר אלפא המימנים הסמוכים לקשר מצביעים לאותו הכוון, בקשרי בטא הם מצביעים לכוונים מנוגדים, הגוף יודע לפרק קשרי אלפא יותר טוב מבטא ברב סוכרים ולכן קשר לו לפרק תאית (צלולוזה) שהיא רב סוכר המצוי בסיבי הצומח.
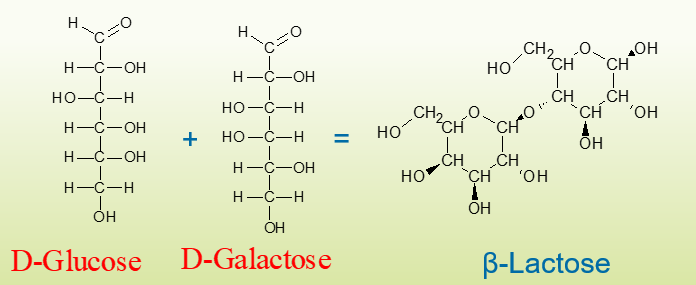
באיור המוצג להלן מולקולת דו סוכר בטא לקטוז אשר מורכבת מחד סוכרים גלקטוז + וגלוקוז. באיור לאחריו גלוקוז+ פרוקטוז= סוכרוז
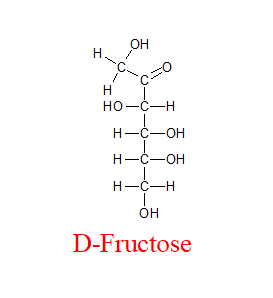
מולקולת הפרוקטוז היא קלטוז (החמצן עם הקשר הכפול לא נמצא בקצה)
דו סוכר של גלוקוז יוצר מלטוז, רב סוכר של גלוקוז יוצר עמילן אם הוא ממקור צמחי, אם הוא ממקור אנדוגני גליקוגן ומכיל גם קצת חלבון.
ליפידים
הנוסחה של חומצת שומן כאשר N=מספר, היא COOH+H3+N(CH2)
הקצה COOH נקרא קרבוקסיל משום שהוא מכיל הידרוקסיל וקרבוניל אז חיברו את השמות.

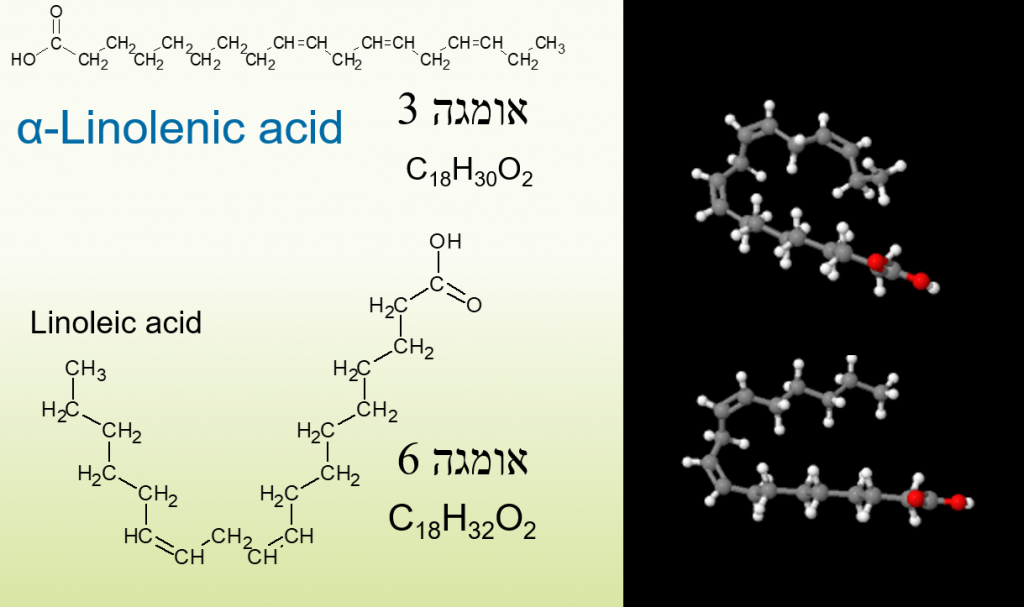
מה שמייחד מולקולות שומן זה מיקומם של קשרים כפולים בין הפחמנים, ומספרם, קשרים אלו מובילים לניתוק קשרי מימן מה שיוצר מבנה שניוני מפוטל למולקולה ומסייע לגוף לפרק אותה. סופרים את מיקום הקשר הכפול הראשון מהצד אשר רחוק מן הקרבוניל
רוב חומצות השומן מצויות במבנה הקרוי טריגליצרידים, 3 חומצות מחוברות לגליצרין/גליצרול
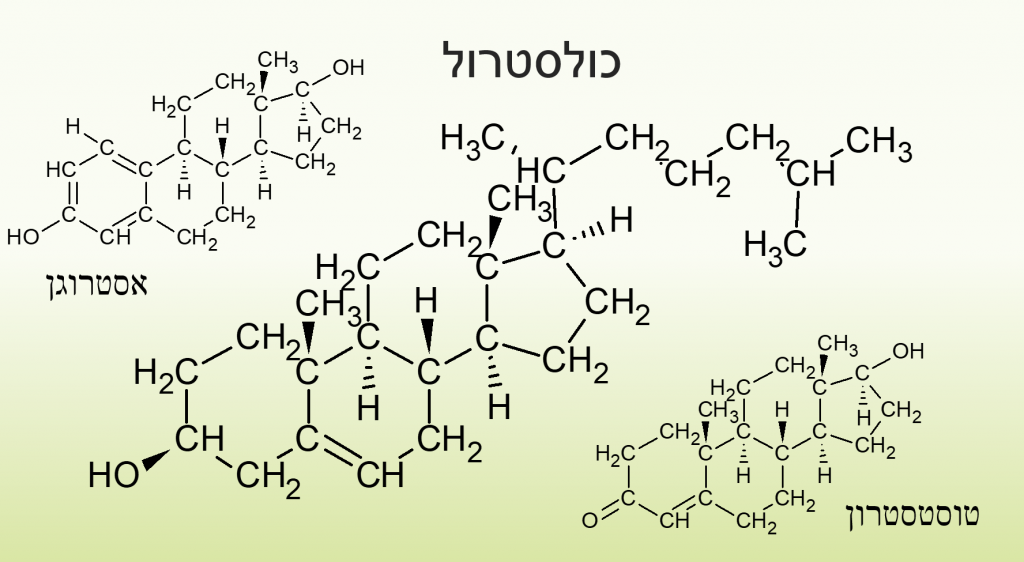
נוסף על כך ישנו שומן ממקור אנדוגני אשר נוצר בכבד ונקרא כולסטרול, כמותו בגוף מזערית אבל יש לו תפקיד בבניין הורמונים סטרואידיים, רמתו בדם משפיע על ריכוז הLDL, VLDL, והHDL שהם שומנים אשר מחוברים לחלבונים אשר פונים לחלק החיצוני ומאפשרים להם לנוע בסביבה מימית ולגוף לזהותם לצורך ספיגה או משוב על רמתם בדם. כילומקרונים מכילים יותר טריגליצרידים.
כולסטרול וטרפנטינים וספינגו ליפידים הם שומנים הנחשבים פשוטים משום שאין להם נוסחה אך נראה שהם מורכבים, חומצות שומן וטריגליצרידים הם שומנים מורכבים על פי ההגדרה.
האיור המוצג לפניכם ניתן לראות שההורמונים הסטרואידים מופקים מכולסטרול
חלבון
חלבון מכיל את הנוסחה המוצגת באיור, COOH+C+NH2+H+R פחמן מרכזי פחמן שמהווה חלק מקצה קרבוקסילי, מצדו האחד קבוצת אמין (חנקן ושני מימנים) ומצדו השני מימן, מתחת ישנו זנב שיכול להשתנות ולשנות את תכונות החלבון

יש לחומצות האמיניות תכונות שונות אשר נקבעות על פי הזנב. אלו המחילות גופרית יכולות ליצור קשר דיסולפידי, אלו המכילות מטען חיובי או שלילי יכולות ליצור ביניהם קשר יוני, חומצות שאינן קוטביות יכולות ליצור קשרי וונדרוולס, ככל שחומצות מכילות יותר O או OH ופחות H הן יותר הידרופיליות וההפך. כל החומצות יכולות ליצור ביניהן קשרים פפטידים/אמינים בין הפחמן לחנקן.
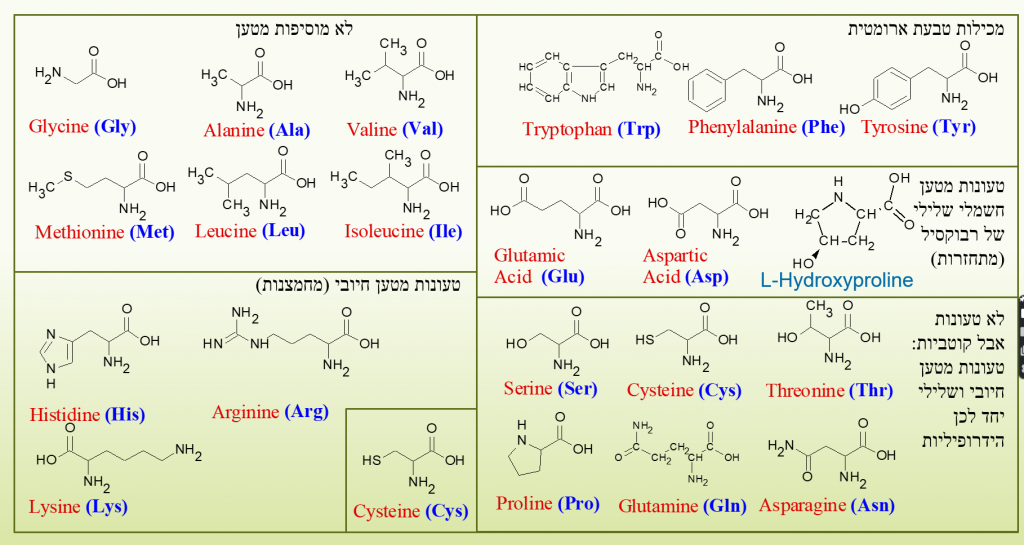
בין החומצות ישנן 20 אשר ניתן לייצר על ידי תהליך השעתוק בגופם של אורגניזמים בכדור הארץ, 9 מתוכן נחשבות חיוניות משום שלא ניתן להחליף אותן ב11 האחרות, לפני כמה שנים היסטדין נחשבה חומצה חיונית למחצה ולכן בספרים ובאיור הזה שהכנתי לפני העדכון יש 8 אבל הדבר התעדכן.

חומצות גרעין
עוד משפחה של חומרים היא משפחת הנוקליאוטידים (חומצות גרעין) הן משמשות לנשאים, אדנין היא חומצת גרעין אשר משמשת לבניין הATP ,כשרמת האדנין עולה הלב פועם יותר לאט, ומשמשות לבניין DNA
אדנין וגואנין שייכים למשפחת הפורינים, הם הבסיסים הגדולים, ותימין וציטוזין שייכים למשפחת הפירימידינים – הבסיסים הקטנים. ריבוז הופך לדיאוקסי ריבוז כאשר הוא מוותר על חמצן

ATP- מורכב מריבוז, שרשרת של מולקולות פוספט (זרחן מוקף בארבע מולקולות חמצן) וחומצת אזנין, הצירוף של אדנין וריבוז נקרא אדנוזין, ומכאן שמו אדנוזיטריפוספט.

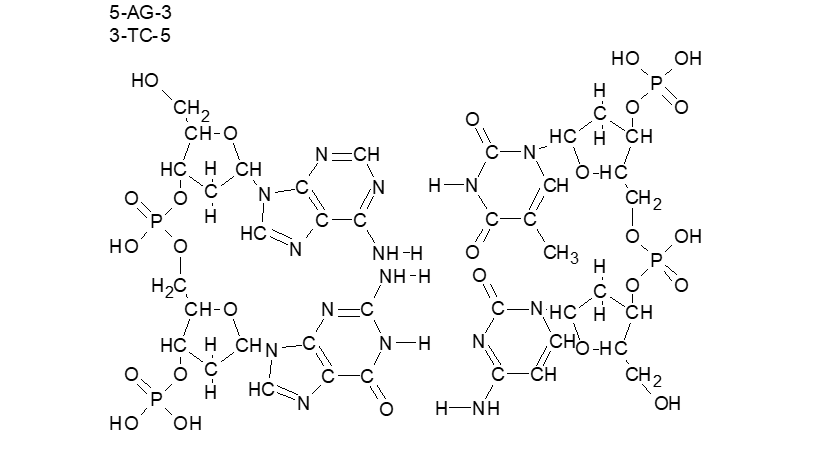
| מבנה DNA |
| מולקולת ה-DNA היא פולימר סלילי הבנוי מיחידה חוזרת. רוחבו של הסליל ואורכה של היחידה החוזרת קבוע. המולקולה היא שרשרת ארוכה, שחוליותיה הם נוקליאוטידים מארבעה טיפוסים, כל נוקליאוטיד מכיל יחידה של סוכר מסוג דאוקסיריבוז, פוספט ובסיס חנקני. ארבעה סוגי בסיסים קובעים את ארבעת טיפוסי הנוקליאוטידים. מספר יחידות אדנין במולקולה זו שווה תמיד לזה של תימין, וזה של גואנין שווה תמיד לזה של ציטוזין. על פי תיאורית הסליל הכפול DNA היא מבנה דמוי סולם. שני מוטות האורך בסולם בנויים כל אחד כשרשרת של שני מרכיבים המתחלפים ביניהם: סוכר – פוספט – סוכר – פוספט וכן הלאה. אל צידו של כל סוכר קשור בסיס חנקני. כאשר שתי השרשראות ניצבות זו מול זו פונים הבסיסים החנקניים שבשרשרת האחת לעבר הבסיסים שבשרשרת האחרת. קישור בין בסיסים משתי השרשרות יוצר זוגות של בסיסים המהווים את שלבי הסולם. שלבי הסולם מורכבים מ2 סוגים של צירופי בסיסים חנקניים: אדנין יחד עם תימין, וגואנין עם ציטוזין. כאמור, אדנין וגואנין שייכים למשפחת הפורינים, הם הבסיסים הגדולים, ותימין וציטוזין שייכים למשפחת הפירימידינים – הבסיסים הקטנים. בין כל שני בסיסים קיימים קשרים כימיים חלשים, קשרי מימן. בין אדנין ותימין נוצרים שני קשרי מימן ובין גואנין וציטוזין נוצרים שלושה קשרי מימן, ולפיכך, בין גואנין וציטוזין קשר חזק יותר. |
| מבנה RNA |
| מולקולת ה-RNA היא שרשרת נוקליאוטידים, המורכבים מבסיסים חנקניים, מזרחה ומהסוכר ריבוז. ההבדל בין הפולימרים בא לידי ביטוי במבנה הבסיס החנקני. מולקלות RNA מורכבות מארבעה סוגי בסיסים: אדנין (A), ציטוזין (C), וגואנין (G).ואורציל (U), זאת בשונה ממולקולת DNA שבה יש תימין (T) במקום אורציל. בניגוד למבנה הגדיל הכפול, האופייני בדרך כלל למולקולות DNA, לרוב RNA מצוי בטבע במבנה של גדיל בודד. עם זאת, גם ב-RNA ייתכן זיווג בסיסים, מה שמביא להיווצרות מבנים שניוניים בגדיל. RNA נייד ורב תכליתי משמש בתהליכים בגוף האדם בשיתוף עם DNA שהינו מולקולה יציבה. |
| כמה סוגי RNA שלמדנו: Messenger RNA, Transfer RNA, RNR הריבוזומלי |
משק האנרגיה בגוף תהליכי התחדשות ושימוש, ותוצרי הפירוק
לאחר שאנו מכירים את המולקולות ניתן ללמוד על הדינאמיקה שלהן בתוך גופנו וחשיבותה ולעבור לנושא הבא והוא תהליכים מטבוליים.
לפני שנכנסים לפירוט אפשר לנסות לפשט התהליכים המטבוליים בקצרה, ניתן למיין כמה תהליכים בגוף לפי הקטגוריות של תהליכי פירוק ותהליכי בניה. זה יכול לסייע בהבנת משק האנרגיה בגוף משום שפירוק מולקולה למולקולה של פחמימה, חלבון או ליפיד למולקולה יותר יציבה (בתנאי קיצון) (מים וCO2) משחרר אנרגיה לסביבה, (האנרגיה הדרושה לייצוב המולקולות לא נדרשת יותר כשהן מומרות למולקולות יציבות), אלו תהליכים שמאפשרים להפיק אנרגיה. לעומתם ישנם תהליכים של הפיכת מולקולות פשוטות למולקולות פחות יציבות בתנאי קיצון אך יציבון בגופנו שניתן לנצל יותר מאוחר.
תהליכים קטבוליים של פירוק: גליקוליזה, מעגל קרבס, שרשרת פירוק אלקטרונים. תהליכים אנבולים של בניה: פוספורלציה (זרחון של אדנוזין) , גלוקונאוגנזה. תהליך הגליקוליזה הפוך מתהליך הגלוקואוגנזה בכדי לוודא שהתהליך לא יתרחש בגוף בו זמנית השני הכוונים יש בשלב מסוים ריאקציה בלטי הפיכה, לכן צריך לעקוף אותה ולהשקיע אנרגיה, הכנת תרשים של התהליך ואיתור השלב של אותה ריאקציה מאפשר להבין למה התהליך מתרחש בכוון מסוים.
הציר המרכזי של תהליכים אלו של פירוק הגלוקוז, תחילה בגליקוליזה אשר לא דורשת חמצן, לאחר מכן במעגל חומצת לימון אשר דורש חמצן ויכול להתרחש במיטוכונדריה, ללא חמצן מולקולות הפירובט גורמות לעליה בחומציות הדם ולכן נהפכות ללקטט אשר יכול לעבור תהליך של גלוקונאוגנזה בכבד אליו דרוש חמצן. הגלוקוז מסיס במים ולכן ישנו תהליך של גליקונאוגנזה (נכתב דומה לגלוקונאוגנזה אבל מתייחס לגליקוגן ולא לגלוקוז) תהליך של יצירת רב סוכר ענק שנאגר בשרירים ובכבד, השרירים חייבים לנצל אותו כי אין להם אנזימים לפירוקו לגלוקוז ללא שימוש להפקת אנרגיה והכבד שולח אותו למחזור הדם כאשר רמת הסוכר בערך נמוך, פירוק שומן במסלול בטא אוקסידציה מהווה גיבוי למצב של חוסר בסוכר, החלבונים משמשים בעיקר לבניה ועוברים מחזור אך השימוש שלהם לצורך אנרגיה מאלץ פליטת חנקן, החנקן מצוי בתור אמוניה שזהו חומר מזיק, לכן ישנו מעגל אמוניה אוריאה שהופך את האמוניה לחומר יותר יציב שניתן לשנע אל הכליות לצורך פינוי כיליתי.
גליקוליזה
תהליך הגליקוליזה של פירוק גלוקוז ל2 מולקולות פירובט, מתרחש בתוך הציטופלזמה של התא, גם כדוריות דם שלא מכילות מיטוכונדירה יכולות לנצלו הוא מאפשר להפיק 2 מולקולות ATP
פירוט שלבי תהליך הגליקוליזה
| שלבי ההשקעה | |
| 1 | גלוקוז מקבל קבוצת זרחה מ-ATP והופך לגלוקוז-6-פוספט. בשל איבוד הזרחה יהפוך ה-ATP ל-ADP. האנזים המזרז את התגובה הוא הקסוקינאז. בנוסף, בכבד קיים גם האנזים גלוקוקינאז שמזרז את התהליך בריכוזי גלוקוז-6-פוספט גבוהים. |
| 2 | גלוקוז 6-פוספט משנה את מבנהו המרחבי, מטבעת של 6 אטומי פחמן לטבעת של 5 פחמנים, והופך לפרוקטוז 6-פוספט. זהו צעד קריטי, שכן ללא השינוי האיזומרי לא יכולים להתרחש השלבים הבאים. זהו שלב הפיך לחלוטין. האנזים המזרז את התגובה הוא פוספוהקסוז איזומרז. |
| 3 | פרוקטוז 6-פוספט מקבל קבוצת זרחה מ-ATP והופך לפרוקטוז 1,6-ביספוספט. זהו שלב ההתחייבות – לאחר שלב זה תמשך הגליקוליזה ללא בקרה משמעותית עד השלב הלפני אחרון. האנזים המזרז את התגובה הוא פוספופרוקטוקינאז (PFK1). |
| 4 | פרוקטוז 1,6-ביספוספט מתפרק לשתי מולקולות תלת-פחמניות: גליצראלדהיד 3-פוספט ודיהידרוקסיאצטון פוספט. האנזים המזרז את התגובה הוא אלדולז. |
| 5 | מולקולת הדיהידרוקסיאצטון פוספט עוברת שינוי מרחבי והופכת ל-גליצראלדהיד 3-פוספט, שנוצרה יחד איתה בשלב 4. עתה קיימות שתי מולקולות של גליצראלדהיד 3-פוספט. האנזים המזרז את התגובה הוא טריוז פוספט איזומראז. |
| שלב הרווח כל השלבים הבאים מתרחשים לכל אחת ממולקולות הגליצראלדהיד 3-פוספט בנפרד – ולכן מתרחשים פעמיים לכל מולקולת גלוקוז. | |
| 6 | כל אחת משתי מולקולות הגליצראלדהיד 3-פוספט מקבלת קבוצת זרחה חופשית (לא מ-ATP) והופכת ל-3,1-ביספוספוגליצרט. שלב זה כולל גם חיזור (קבלת פרוטונים) של מולקולות +NAD ל-NADH. האנזים המזרז את התהליך הוא גליצראלדהיד 3-פוספט דהידרוגנז. |
| 7 | ביספוספוגליצרט 1,3 מאבד קבוצת זרחה והופך ל-3-פוספוגליצרט. קבוצת הזרחה עוברת למולקולת ADP, שהופכת למולקולת ATP. זהו השלב האקסרוגני הראשון בגליקוליזה – השלב הראשון בו הגוף מפיק אנרגיה מהגליקוליזה. האנזים המזרז את התהליך הוא פוספוגליצרט קינאז. |
| 8 | קבוצת הזרחה שנותרה ב-3-פוספוגליצרט משנה מקום (באמצעות אנזים) ונודדת לאטום הפחמן הסמוך; התרכובת הופכת ל-2-פוספוגליצרט. האנזים המזרז את התהליך הוא פוספוגליצרט מוטאז. |
| 9 | בי פוספוגליצרט מאבד מולקולת מים והופך לפוספואנול פירובט. האנזים המזרז את התהליך הוא אנולאז. |
| 10 | פוספואנול פירובט מאבד קבוצת זרחה והופך לפירובט. קבוצת הזרחה עוברת למולקולת ADP ליצירת ATP. האנזים המזרז את התהליך הוא פירובט קינאז. |
גלוקונאוגנזה-
גלוקונאוגנזה- תהליך חידוש גלוקוז אשר מתרחש ברובו בכבד ובאופן חלקי בכליות, נכתב דומה לגליקוגנזה אבל מתייחס לגלוקוז ולא לגליקוגן, המולקולות אשר מעורבות בתהליך הן: פירובט, לקטט, גליצרול כל חומצות האמינו, מלבד לאוצין וליזין. התהליך חשוב משום שהגלוקוז דרוש להזנת תאי עצב, וכדוריות דם אדומות ולכן הגוף מחדש אותו גם אם זה כרוך בפירוק חלבון, התהליך צורך 6 מולקולות ATP ומחזיר 2 מתוכן כך שהוא צורך 4, שלבי התהליך:
| 1 | המרת מולקולות לפירובט |
| 2 | תהליך הפוך מתהליך הגליקוליזה, חלק מן הראקציות של הגליקוליזה הפיכות וחלק לא ניתן להפעיל בכוון הפוך (ריאקציות של ביקוע המולקולה ל2) ולכן צריך מסלול עוקף בכדי להפוך 2 מולקולות פירובט למולקולת גלוקוז. |
מעגל קרבס (מעגל חומצת לימון/חומצה ציטרית)
מעגל קרבס מכונה מעגל משום שיש בו מולקולה של חומצת לימון אשר מתחברת למולקולת פירובט, מולקולת הפירובט מתפרקת לCO2 ולמימן, עד פירוקה הסופי ואז נותרת מולקולת חומצת הלימון כפי שהייתה לפני התהליך מוכנה לסיבוב נוסף. יש בתהליך השקעה של 2 מולקולות ATP אבל רווח אנרגטי של 4 מולקולות ATP ישירות, ועוד 28 שיבצרו על ידיNADH+, FAHH2+ i , אצל חיידקים ניתן להפיק גם 36 מולקולות, הסרטון המצורף על חיידקים ולכן המספרים שם שונים, להלן פירוט של התהליך וכל האנזימים המעורבים.
פירוט שלבי מעגל קרבס
| 1 | אוקסאלואצטט מתחבר לאצטיל קואנזים A ויוצר ציטרט (Citrate), מולקולה בת שישה פחמנים. שלב זה מתרחש בעזרת האנזים citrate synthase. זהו שלב חד כיווני. |
| 2 | ציטרט עובר שינוי איזומרי והופך לאיזוציטרט (Isocitrate). שלב זה מתרחש בעזרת האנזים aconitase. |
| 3 | איזוציטרט מאבד פחמן דו-חמצני והופך לאלפא-קטוגלוטרט (α-Ketoglutarate), מולקולה בת חמישה פחמנים. שלב זה מתרחש בעזרת האנזים isocitrate dehydrogenase. |
| 4 | אלפא-קטוגלוטרט מאבד פחמן דו-חמצני, מתחבר לקואנזים A ויוצר סוקציניל (Succinyl) קואנזים A. שלב זה מתרחש בעזרת האנזים α-Ketoglutarate dehydrogenase. כעת נותרו 4 פחמנים, אם כן ניתן לראות כי שני אטומי פחמן נכנסים למעגל כיחידות acetyl ושני אטומי פחמן עוזבים אותו בצורת פחמן דו-חמצני. |
| 5 | סוקציניל קואנזים A מאבד קואנזים A והופך לסוקצינט (Succinate). שלב זה מתרחש בעזרת האנזים succinyl coA synthase בתהליך היוצר מולקולת GTP. |
| 6 | סוקצינט הופך לפומרט (Fumarate). שלב זה מתרחש בעזרת האנזים Succinate dehydrogenase. |
| 7 | פומרט הופך למלט (Malate). שלב זה מתרחש בעזרת האנזים fumarase. |
| 8 | מלט הופך לאוקסאלואצטט. שלב זה מתרחש בעזרת האנזים malate dehydrogenase |
גליקוגנוליזה
גליקוגנוליזה- פירוק הגליקוגן ושחרורו לדם מהכבד/ צריכתו בשרירים על ידי גליקוליזה.
ישנם מאגרי גליקוגן בשרירים ובכבד, זהו רב סוכר
| גליקוגנוליזה (מיוונית, ליזיס = פירוק) היא התהליך הביוכימי של פירוק גליקוגן, על ידי הסרת יחידה של גלוקוז מזורחן, ליצירת גלוקוז-1-פוספט ההופך לגלוקוז-6-פוספט, תוצר ביניים של הגליקוליזה. |
| גליקוגנוליזה מתרחשת ברקמות שריר ובכבד, שם יש מאגרי גליקוגן, כתוצאה מגירוי הורמונלי של אפינפרין או גלוקגון (פפטיד המשתחרר מהלבלב כתגובה לרמות גלוקוז נמוכות בתאי אלפא של איי לנגרהנס). |
| תאי כבד מסוגלים לפרק את הגלוקוז-6-פוספט בגליקוליזה או להסיר את קבוצת הפוספט בעזרת האנזים גלוקוז-6-פוספטאז ולשחרר את הגלוקוז החופשי לזרם הדם משם ינוצל על ידי תאים אחרים. |
| תאי שריר אנושיים אינם כוללים גלוקוז-6-פוספטאז וממילא לא ישחררו את הגלוקוז אלא ינצלו אותו בתהליך גליקוליזה. |
גליקוגנזה
תהליך חידוש הגלוקוז, אנזימים מסויימים משתתפים גם בגליקוליזה וגם בגליקוגנזה, ואנזימים מסויימים ייחודיים למסלול מטבולי אחד מהם, הורמונים מאפשרים בקרה על ידי הפעלת אותם האנזימים. בכדי להבין מה כוון התהליך יש לתאר אותו בטבלה:
| גליקוגנזה | |
| 1 | גלוקוז הופך לגלוקוז-6-פוספט על ידי גלוקוקינאז או הקסוקינאז. |
| 2 | גלוקוז-6-פוספט הופך לגלוקוז-1-פוספט על ידי פוספו-גלוקו-מוטאז, דרך תוצר הביניים, גלוקוז-6,1-פוספט. |
| 3 | גלוקוז-1-פוספט הופך ל-UDP-גלוקוז על ידי יורידיל-טרנפראז היוצר גם פירופוספט העובר הידרוליזה על ידי פירופוספטאז ליצור 2 מולקולות זרחן. |
| 4 | מולקולות גלוקוז נאספות בשרשרת על ידי גליקוגן-סינטאז, הדורש קיום שרשרת תחל ראשונית (נוצרת על ידי גליקוגנין). |
| 5 | סיעופים נוצרים על ידי אנזים סיעוף [amylo-α(1:4)->α(1:6)transglycosylase] המעביר את סוף השרשרת לאמצע בקשר אלפא-1:6, משם השרשרת ממשיכה לגדול בקשרי 1:4 |
בקרה על משק הגליקוגן (גליקוגנזה או גליקוגנוליזה)
הבקרה נעשית בשתי דרכים, בקרה אלוסטרית- שינוי במבנה חלבונים כתגובה לגלוקוז6 פוספט/ ATP שמעודדים הפסקת פירוק גליקוגן (גליקוגנוליזה) ושינויים אשר מעודדים פירוק גליקוגן בתגובה לAMP. בקרה הורמונאלית של הורמונים אשר מעודדים פירוק : GH, קורטיזול, אדרנלין וגלוקגון, והורמונים אשר מעודדים בניה: אינסולין ועוד.
ATP סינתאז
F0– אזור ממברנלי, דרכו עוברים פרוטונים המגיעים מהחלל הבין־ממברנלי ועוברים את המטריקס. F1-אזור פריפריאלי, באזור זה נכנסות מולקולות ADP + Pi ומתקבלת מולקולת ATP.
אזור F1 מורכב משלוש תתי־יחידות של אלפא ובתא, אל תת-היחידה הראשונה נכנסים ADP + Pi, נוצרת מולקולת ATP שעוברת אל תת-היחידה השנייה ויוצאת דרך תת-היחידה השלישית. המעבר בין תת-היחידות מתרחש הודות למעבר הפרוטונים דרך החלבון F0, אשר גורם לסיבוב ובכך לשינוי קונפורמציה בחלק F1 של החלבון ולשחרור מולקולת ה־ATP. האנרגיה המושקעת התהליך משמשת להוצאת ה־ATP מהחלבון ולא על מנת ליצור את ה־ATP. יצירת ה־ATP לא דורשת השקעה של אנרגיה.
בסה"כ נדרשים 4 פרוטונים על מנת ליצור מולקולת ATP אחת: 3 פרוטונים כדי ליצור בפועל את ה ATP ועוד פרוטון כדי להעביר ADP + Pi אל תוך המטריקס.
משק השומן
גליקוליזה- השומן נאגר ברובו כטריגליצרידים, יכול להתפרק בציטופלזמה בתהליך של גליקוליזה כאשר הוא מתפרק לרכיבים, חומצות השומן ימשיכו לשלב נוסף והגליצרול יכול לחזור לכבד להפוך לפירובאט, ומשם להכנס למעגל קרבס לצורך ייצור אנרגיה, או לגלוקונאוגנזה לשמירת רמת הגלוקוז בדם.
β-אוקסידציה במיטוכונדריה – פירוק חומצות שומן ליחידות דו-פחמניות, אצטיל-CoA ,הנכנסות למעגל קרבס להפקת ATP .גם מהפירוק עצמו נוצרים אקויוולנטים מחזרים, המומרים ל-ATP. קטוגנזיס – כאשר יש חוסר במקורות אנרגיה חיצוניים, יש פירוק מוגבר של חומצות שומן, המביא לעודף של אצטיל-CoA . מצב זה מוביל את הגוף לייצור גופי קטו.
ליפוגנזה – יצירת חומצות שומן מאצטיל-CoA במצב של עודף אנרגיה. זהו תהליך הדורש אנרגיה )ATP.)
ליפוליזה
הוא המסלול המטבולי שבאמצעותו הידרוליזה טריגליצרידים בשומנים לגליצרול ושלוש חומצות שומן. הוא משמש לגיוס אנרגיה מאוחסנת במהלך צום או פעילות גופנית, ליפוליזה נגרמת על ידי מספר הורמונים, בהם גלוקגון, אפינפרין, נוראדרנלין, הורמון גדילה, קורטיזול ועוד. בגוף מכונים מאגרי שומן כרקמת שומן. באזורים אלה הטריגליצרידים התאים (כלומר בתוך התא) מאוחסנים בטיפות ליפידים ציטופלסמטיות. כאשר ליפאזים זרחניים הם יכולים לגשת לטיפות ליפידים ובאמצעות שלבים מרובים של הידרוליזה.
| 1 | פירוק הטריגליצרידים לחומצות שומן וגליצרול. כל שלב של הידרוליזה מוביל להסרת חומצת שומן אחת. השלב הראשון והצעד המגביל את הקצב של הליפוליזה מתבצע על ידי טרגליצרידים ליפאז (Adipose triglyceride lipase). אנזים זה מזרז את ההידרוליזה של triacylglycerol ל diacylglycerol. |
| 2 | לאחר מכן, ליפאז רגיש הורמונלי (HSL) מזרז את ההידרוליזה של דיאקיל-גליצרול למונואקיל-גליצרול ומונואקיל-גליצרול-ליפאז (MGL) |
| 3 | מזרז את ההידרוליזה של מונואציל-גליצרול לגליצרול. |
חמצון בטא (β oxidation)
כאשר משק הפחמימות עני, אין מספיק גלוקוז הגוף משקיע אנרגיה בחידוש המלאי, המקור הבלעדי לחידוש מלאי הסוכר בגוף הוא השומן, ולכן יש צורך בפירוק שומן.
התהליך- חמצון בטא מצריך פירוק שומן הובלתו אל התאים ושם הוא עובר ארבעה תהליכים עוקבים המתבצעים על חומצת שומן מסוימת אשר בסופם מתקבלת מולקולת אצטיל קואנזים A חופשית המועברת ישירות אל מעגל קרבס, פירוט השלבים מוצג בטבלה:
| שני שלבי הכנה ו4 שלבים של התהליך | |
| 1 | פירוק הטריגליצריד למרכיביו, ושינוע חומצות השומן החופשיות לתאי הגוף השונים. גם ניצול חומצות השומן מהמזון דורש מספר שלבים מקדימים הכוללים את אריזת הטריגליצרידים בליפופרוטאינים מיוחדים. פירוק מאגרי השומן- תהליך פירוק הטריגליצרידים בתאי השומן מתבצע הודות לעלייה בהורמוני הגלוקגון או האפינפרין בדם. הורמונים אלה גורמים בראש ובראשונה להפעלת אנזים ליפאז ספצפי (Hormone Sensitive Lipase) אשר מתחיל לפרק את הטריגליצריד למרכיביו. חומצות השומן החופשיות משונעות בשלב הבא, על ידי חלבון האלבומין אל תאי הגוף השונים. בהגיעם אל התאים, מסוגלות חומצות השומן לעבור את ממברנות התאים הודות לשומניות שלהם ולמסיסות הנמוכה שלהם במים. |
| 2 | המעבר למיטוכונדריה- בעוברם את קרום התא מגיעות חומצות השומן לציטוזול התאי ומיד מגיבות עם קואנזים A ויוצרות מולקולת אציל קואנזים A אשר נחוצה לתהליך חמצון בטא. ריאקציה זו דורשת השקעה של אנרגיה ולכן מולקולת ATP אחת מנוצלת בתהליך. אולם מולקולות האציל קואנזים A אינן מסוגלות להיכנס אל תוך פנים המיטוכונדריה, מפני שהן אינן מצליחות לחדור את הקרום הפנימי שלהן. המעבר למיטוכנדריה מתאפשר אם כך הודות למולקולת הקרניטין, אשר מגיבה עם האציל קואנזים A ליצירת מולקולת אציל קרניטין. ריאקציה זו מזורזרת בנוכחות האנזים קרניטין אציל טְרַנְסְפֵרַאז I. אציל קרניטין חודרת את הקרום הפנימי ובתוך חלל המיטוכונדריה חוזרת לצורת אציל קואנזים A בנוכחות האנזים אציל טרנספראז II. |
| כעת מולקולות האציל קואנזים A מתחילות את חמצון הבטא בפועל. תהליך החמצון- תהליך חמצון הבטא של חומצת השומן בעלת 16 פחמנים פלמיטט לחומצת שומן בעלת 14 פחמנים חומצה מיריסטית ואצטיל קואנזים A. כאמור, מטרת העל של התהליך הזה היא לבצע חיתוכים של מולקולות אצטיל קואנזים A על מנת לנצל אותן במעגל קרבס לצורך הפקת אנרגיה. לתהליך 4 שלבים: | |
| A | סילוק זוג מימנים מחומצת השומן ויצירת קשר כפול זמני מסוג טראנס בין פחמן האלפא לבין פחמן הבטא. ריאקציה זו מתרחשת הודות לאנזים אציל קואנזים A דֵהִידְרוֹגֵנָאז והמטבוליט שמתקבל נקרא טְרַאנְס-Δ אֶנוֹאִיל קואנזים A. אנזים זה נעזר במולקולה נשאית האלקטרונים FAD המשמשת לו כקואנזים. זהו השלב החשוב ביותר אשר קובע את קצב התהליך כולו. |
| B | הכנסת מולקולת מים באמצעות האנזים אֶנוֹאִיל הִידְרַטָאז, דבר הגורם לפתיחת הקשר הכפול וקבלת מולקולת הִידְרוֹקְסִיאָצִיל קואנזים A. |
| C | הוצאת מימנים נוספת, הפעם בעזרת מולקולה נשאית האלקטרונים NAD והאנזים בטא הידרוקסיאציל קואנזים A דהידרוגנאז לקבלת המטבוליט קֵטוֹאִצִיל קואנזים A. |
| D | חיתוך מולקולת קואנזים A בעזרת האנזים תִּיאוֹלִאז מתוך מולקולת קטואציל קואנזים A. דבר הגורם לחומצת השומן להקצר בשני פחמנים. אצטיל קואנזים A מנוצל במעגל קרבס בעוד שחומצת השומן הנותרת חוזרת על אותו התהליך מהתחלה כאשר היא נדחסת עם קואנזים A, לקבלת אציל קואנזים A חדשה בנוכחות אותו התיאולאז |
מעגל פירוק אוריאה
| מעגל האוריאה מורכב משני שלבים עיקריים: השלב המיטוכונדריאלי והשלב הציטוזולי. השלב המיטוכונדריאלי מורכב משתי ריאקציות בעוד שהציטוזולי משלוש. אולם היות שביונקים מתרחש מעגל האוריאה רק בתאי הכבד יש צורך במספר שלבים מקדימים על מנת להבטיח את שינוע האמוניה מתאי הגוף השונים אל הכבד. השינוע עצמו מתבצע על ידי שתי חומצות האמיניות: גלוטמין ואלנין כשהשינוע על ידי הגלוטמין מתרחש מכל תאי הגוף והשימוש באלנין מתרחש ספציפית רק מתאי השריר. | |
| 1 | השלב המקדים הראשון כולל את פירוק קבוצות האמינו (בצורת יון האמוניום) מחומצות האמינו (שמתקבלות מצריכת חלבונים במזון או בפירוק רקמות המכילות חלבונים בגוף כגון רקמת שריר וחלבוני הפלזמה) ושתילתן במולקולות של אלפא קטו גלוטרט או פירובט (בתאי השריר בלבד). |
| בתאי הגוף מתרחש התהליך של יצור חומצה אמינית גלוטמט על ידי שתילה של קבוצות האמינו במולקולה של אלפא קטו גלוטרט על ידי אנזימים ספציפיים הנקראים אמינוטרספראז המותאמים כל אחד לחומצת אמינו שלו. | |
| בתאי השריר מתרחשת ריאקציה שונה, שבה לאחר יצור הגלוטמט, מועברת הקבוצה האמינית שלה למולקולה של פירובט ההופכת אותה לאלנין. ריאקציה זו מזורזת על ידי האנזים אלנין אמינוטרנספראז. מולקולת האלנין מסוגלת כעת לנדוד בזרם הדם אל עבר הכבד, אולם מולקולת הגלוטמט שנוצרה אינה נודדת לכבד לפני הפיכתה לגלוטמין, מפני שיש עדיפות לשנע גלוטמין אשר מורכבת משתי קבוצות אמינו על פני גלוטמט בעלת קבוצה אחת בלבד (יתרון הכמות של האמוניה המסולקת). | |
| 2 | שלב מקדים שני |
| השלב המקדים השני כולל את הפיכת מולקולת הגלוטמט לגלוטמין, שינועה לעבר הכבד והפיכתה מחדש למולקולת גלוטמט באתר המטרה. בריאקציה הראשונה בעצם "מוקרבת" מולקולת גלוטמט אחרת שנוצרה בשלב המקדים הראשון כלומר, נחתכת הקבוצה האמינית שלה על ידי האנזים גלוטמט דהידרוגנאז ומושתלת על מולקולת גלוטמט אחרת. ריאקציה זו מזורזת על ידי האנזים גלוטמין סינתאז והתוצאה היא קבלת הגלוטמין. כעת מולקולת הגלוטמין מסוגלת לנדוד בדומה לאלנין בזרם הדם לכבד, וברגע כניסתה לתאי הכבד הופכת מחדש למולקולת גלוטמט על ידי הריאקציה המתווכת על ידי האנזים גלוטמינאז. התוצאה היא כאמור גלוטמט וקבוצה אמינית חופשית אחת הנכנסת ישירות למעגל האוריאה. | |
| מולקולת האלנין שהגיעה מתאי השריר "מחזירה" את קבוצת האמינו שלה לאלפא קטו גלוטרט ובכך חוזרת להיות פירובאט חופשי שהופך לגלוקוז בתהליך הגלוקונאוגנזה. האלפא קטו גלוטרא הופך, בקבלת קבוצת האמין, לגלוטמאט שנכנס לשלב המיטוכונדריאלי של מעגל האוריאה. | |
| 3 | השלב המיטוכונדריאלי |
| זהו השלב הראשון במעגל האוריאה עצמו והוא מתרחש במיטוכונדריות של תאי הכבד. | |
| לגלוטמט אשר התקבל מהגלוטמין יש כעת שתי אפשרויות: או באמצעות האנזים, שכבר השתתף בשלב המקדים השני – גלוטמט דהידרוגנאז לוותר על קבוצת האמינו שנותרה לו או להדחס יחד עם מולקולת אוקסלואצטט אשר מקורה במעגל קרבס ולהרכיב חומצת אמינו אספרטט שתכנס למעגל האוריאה בשלב מאוחר יותר. | |
| הקבוצה האמינית שמתקבלת מהאפשרות הראשונה עוברת שלב של דחיסה עם מולקולת ביקרבונט בנוכחות ATP והאנזים קָרְבּוֹמוֹאִיל פוספט סינתאז I והתוצר שמתקבל היא מולקולת קרבומואיל פוספט אשר נכנסת למעגל האוריאה על ידי התרכבות עם מולקולת האוֹרְנִיטִין שהוא גם המטבוליט האחרון במעגל והתוצר שמתקבל הוא המטבוליט הראשון במעגל – צִיטְרוּלִין. מולקולת הציטרולין מוצאת אל מחוץ למיטוכונדריה אל עבר הציטוזול על ידי חלבון נשא מתאים. | |
| 4 | השלב הציטוזולי |
| בציטוזול של תאי הכבד על ידי הוספת מולקולת ATP וחומצת האמינו אספרטט (שהתקבלה מדחיסת גלוטמט עם האוקסלואצטט) לציטרולין נוצרת מולקולת אַרְגִּינִינוֹסוּקְסִינָט. בשלב הבא עוזבת מולקולת הפומרט את מעגל האוריאה ומועברת אל מעגל קרבס, כתוצאה מזה הופך הארגינינוסוקסינט לחומצת אמינו ארגינין טהורה. לאחר מכן בנוכחות האנזים ארגינאז משתחררת האוריאה שמועברת בזרם הדם לעבר הכליות והתוצר שמתקבל במעגל הוא האורניטין מחדש שחוזר בעזרת נשא מתאים חזרה אל המיטוכונדיה ומאפשר התחלת המסלול מחדש. |